While humans have observed the heritability of traits for millennia (it is the basis of agriculture), the earliest experiments on understanding inheritance were by the Austrian monk Gregor Mendel. Mendel worked with peas, which as fast growing plants were easy to work with and make observations on. Mendel's observations were first made with simple genetic traits, as a time when there was neither an understanding of evolution by natural selection or molecular genetics, the same principles apply to more complex inheritance patterns.
In mendelian genetics, for a given trait (hair color, eye color, disease etc) in an organism that reproduces sexually, an individual has two versions of the trait, known as alleles. Of these alleles, they inherited one from each parent. Furthermore, when an individual has offspring they only pass on one allele (Figure 1).

Different alleles have properties in relation to one another. Dominant alleles are alleles which if they are present are expressed by the individual. An individual only needs one copy to have the trait associated with the allele. Recessive alleles by contrast are alleles that are only expressed if the dominant allele is missing. This contrast is usually because the dominant allele is for a new function that would not otherwise be present if the allele was not, so the presence of a function is expressed, while the absence of a function is recessive, as if that effect would not be noticeable unless the function was entirely absent. There's dominant and recessive alleles are denoted T and t respectively, with a letter used relevant to the trait.
In a classic punnet square of offspring from two parents who have both dominant and recessive alleles (known as heterozygous) (Figure 2), there are three possible states for offspring, they may have only the dominant allele (homozygous dominant), heterozygous, or only the recessive allele (homozygous recessive). These occur in a 1:2:1 ratio, while the ratio of expressing the trait is 3:1 in favour of the dominant trait.

In the case of a recessive genetic disorder, then two healthy parents may be carriers of a recessive allele for the disease. In this case they have a 1 in 4 chance of having children with the genetic disorder. While 2 in 3 of any unaffected children they have will be carriers. Because people often don't have large families, they may not be aware they carry such a condition.
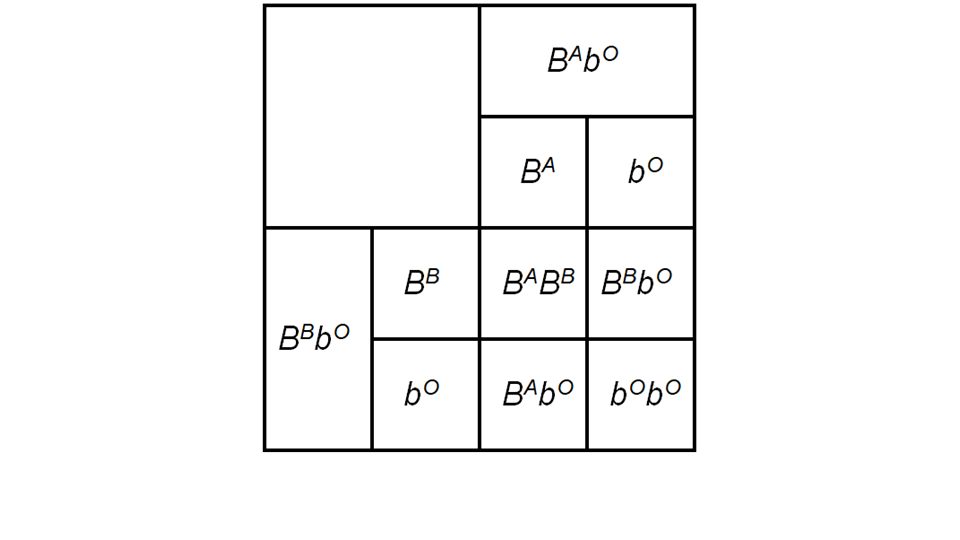
There are also codominant alleles, these are alleles that are equally as dominant as one another. Blood groups are controlled by such codominant alleles, with both the A and B blood group alleles being able to be equally expressed. This means that parents who each have A and B blood groups can have a child with A, B, AB or O blood groups (Figure 3). Cat fur colour is also controlled in a similar way, but this is complicated by the fact that fur colour is expressed on sex chromosomes. Codominant alleles are often described in the form Tt to indicate the codominance, and distinguish the alleles.
If alleles are found on the sex chromosomes (the X or Y chromosomes) then the sex of which parent is the carrier, and the sex of offspring can affect the expression and inheritance of the alleles (Figure 4). Because females have two X chromosomes, alleles are expressed largely in the same way as those on other chromosomes. Males however do not have an additional X chromosome, so if a male has a recessive allele on their X chromosome, it will always be expressed as there is no dominant allele. A classic example of this is in hemophilia, a genetic disease which people inherit from their mothers but that only affects men.

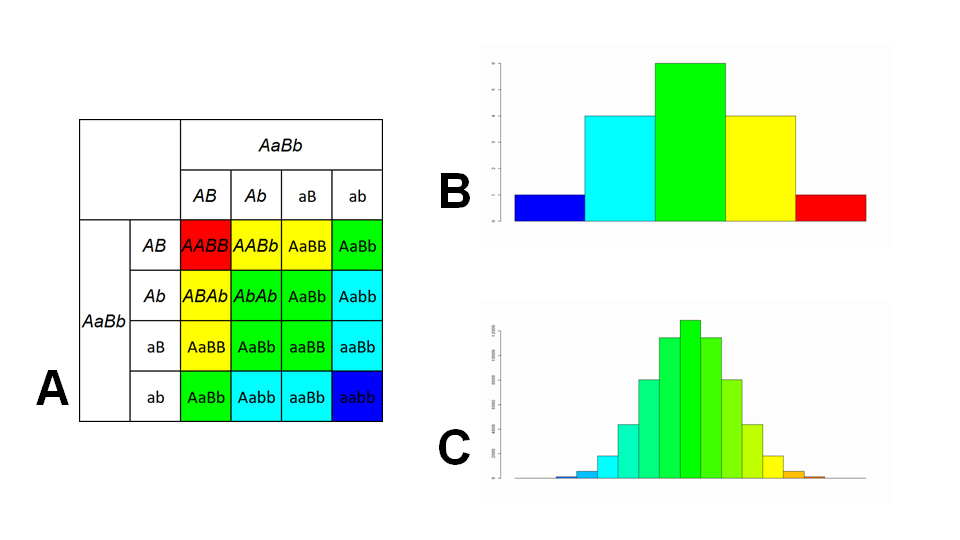
Many traits are not as simple as classical Mendelian genetics involving a single allele. Some traits involve multiple alleles and the number of each allele inherited (especially in codominant traits) creates a range of how much the trait is expressed, which is dependent on how much of each allele is inherited. Like how parents who are heterozygous can have any offspring with any genotype, parents who are in the middle of this range can have offspring who are at both extremes of this range, depending which alleles are inherited by their offspring (Figure 5).
Some traits however are not inherited in a way that conforms to Mendelian genetics. Alleles are not in isolation, and often interact with other genes to express their traits. Furthermore the expression of alleles is also often affected by the individual's environment, and while an allele may give an individual a tendency towards a particular trait, whether this trait is observed will depend on lifestyle factors. However despite these nuances, the inheritance of individual alleles can still be modelled using Mendelian genetics.